Introduction
One of the tenets of modern neuroscience is that the brain modifies the strengths of its synaptic connections (“weights”) during learning in order to better adapt to its environment. However, the underlying learning rules (“weight updates”) in the brain are currently unknown. Many proposals have been suggested, ranging from Hebbian-style mechanisms that seem biologically plausible but are not very effective as learning algorithms in that they prescribe purely local changes to the weights between two neurons that increase only if they activate together -- to backpropagation, which is effective from a learning perspective by assigning credit to neurons along the entire downstream path from outputs to inputs, but has numerous biologically implausible elements.
A major long-term goal of computational neuroscience is to identify which learning rules actually drive learning in the brain. A further difficulty is that we do not even have strong ideas for what needs to be measured in the brain to quantifiably assert that one learning rule is more consistent with those measurements than another learning rule. So how might we approach these issues? We take a simulation-based approach, meaning that experiments are done on artificial neural networks rather than real brains. We train over a thousand artificial neural networks across a wide range of possible learning rule types (conceived of as “optimizers”), system architectures, and tasks, where the ground truth learning rule is known, and quantify the impact of these choices. Our work suggests that recording activities from several hundred neurons, measured semi-regularly during learning, may provide a good basis to identify learning rules -- a testable hypothesis within reach of current neuroscience tools!
Background: A Plethora of Theories and a Paucity of Evidence
The brain modifies the connections between neurons during learning to improve behavior; however, the underlying rules that govern these modifications are unknown. The most famous proposed learning rule is “Hebbian learning”, also known by the mantra: “neurons that fire together; wire together”. In this proposal, a synaptic connection strengthens if one neuron ("pre-synaptic") consistently sends a signal to another neuron ("post-synaptic"). The changes prescribed by Hebbian learning are “local” in that they do not take into account a synapse’s influence further downstream in the network. This locality makes learning rather slow even in the cases where additional issues, such as the weight changes becoming arbitrarily large, are mitigated. Though there have been many suggested theoretical strategies to deal with this problem, commonly involving simulations with artificial neural networks (ANNs), these strategies appear difficult to scale up to solve large-scale tasks such as ImageNet categorization [1].
This property of local changes is in stark contrast to backpropagation, the technique commonly used to optimize artificial neural networks. In backpropagation, as the name might suggest, an error signal is propagated backward along the entire downstream path from the outputs of a model to the inputs of the model. This allows credit to be effectively assigned to every neuron along the path.
Although backpropagation has long been a standard component of deep learning, its plausibility as a biological learning rule (i.e. how the brain modifies the strengths of its synaptic connections) is called into question for several reasons. Chief among them is that backpropagation requires perfect symmetry, whereby the backward error-propagating weights are the transpose of the forward inference weights, for which there is currently little biological support [2, 3].
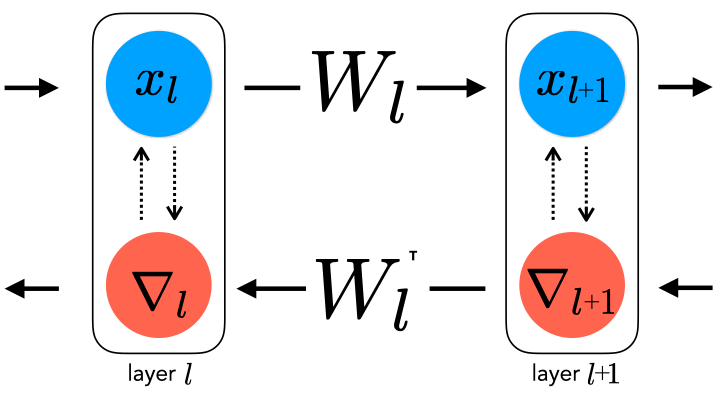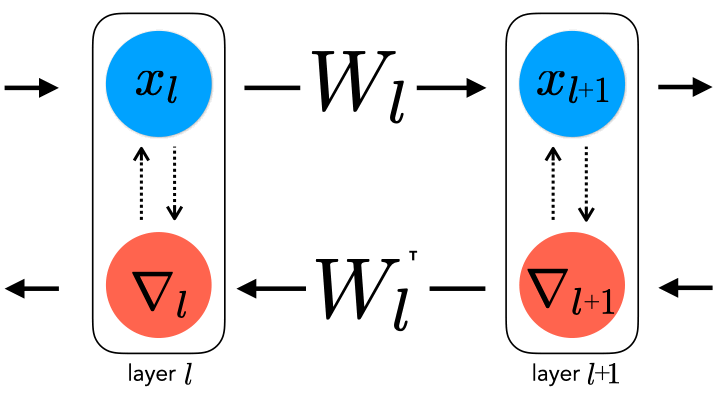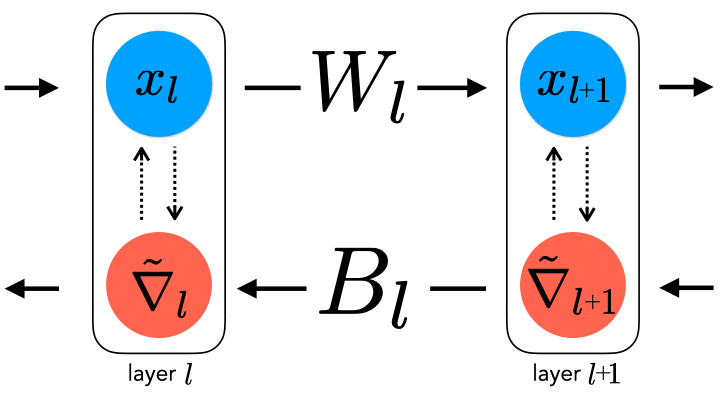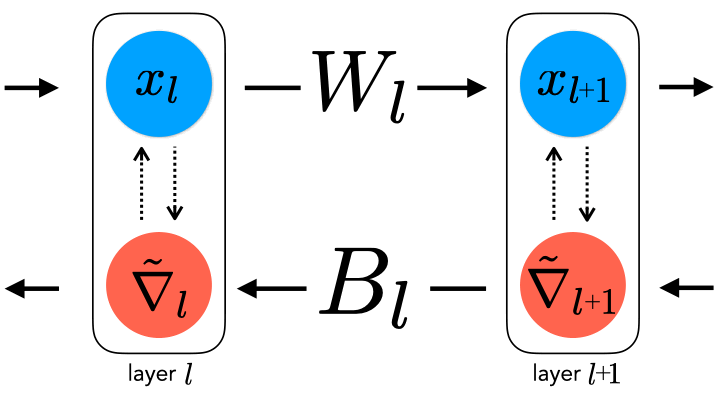
Recent approaches, from us and others [4, 5], introduce approximate backpropagation strategies that do not require this symmetry, and can still succeed at large-scale learning as backpropagation does. However, given the number of proposals, a natural question to ask is how realistic they are. At the moment, our hypotheses are governed by domain knowledge that specifies what “can” and “cannot” be biologically plausible (e.g. “exact weight symmetry is likely not possible” or “separate forward and backward passes during learning seem implausible”), as well as characterizations of ANN task performance under a given learning rule (which is not always directly measurable from animal behavior). In order to be able to successfully answer this question, we need to be able to empirically refute hypotheses. In other words, we would ideally want to know what biological data to collect in order to claim that one hypothesis is more likely than another.
More concretely, we can ask: what specific measurements from the brain, in the form of individual activation patterns over time, synaptic strengths, or paired-neuron input-output relations, would allow one to draw quantitative comparisons of whether the observations are more consistent with one or another specific learning rule? For example, suppose we record neural responses (“activation patterns”) while an animal is learning a task. Would these data be sufficient to enable us to broadly differentiate between learning rule hypotheses, e.g. by reliably indicating that one learning rule’s changes over time more closely match the changes measured from real data than those prescribed by another learning rule?
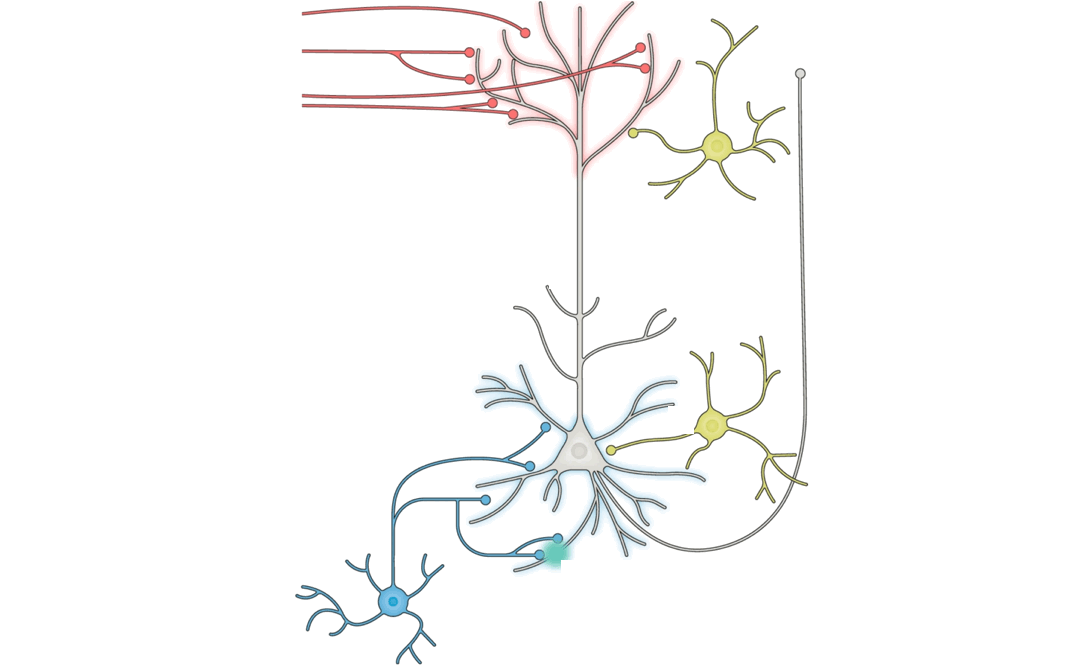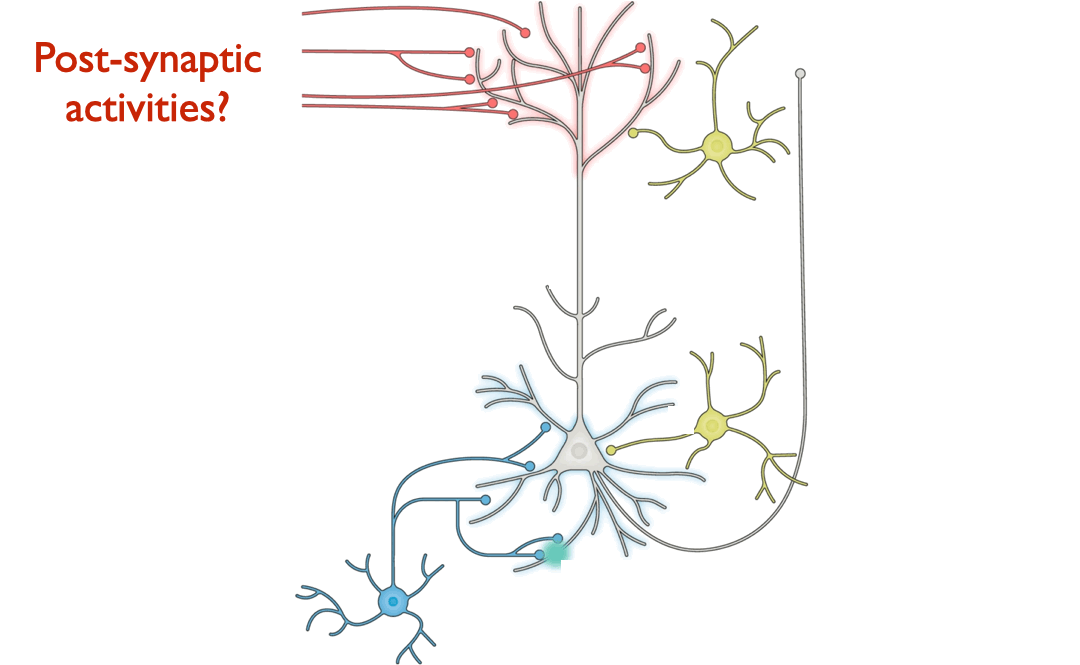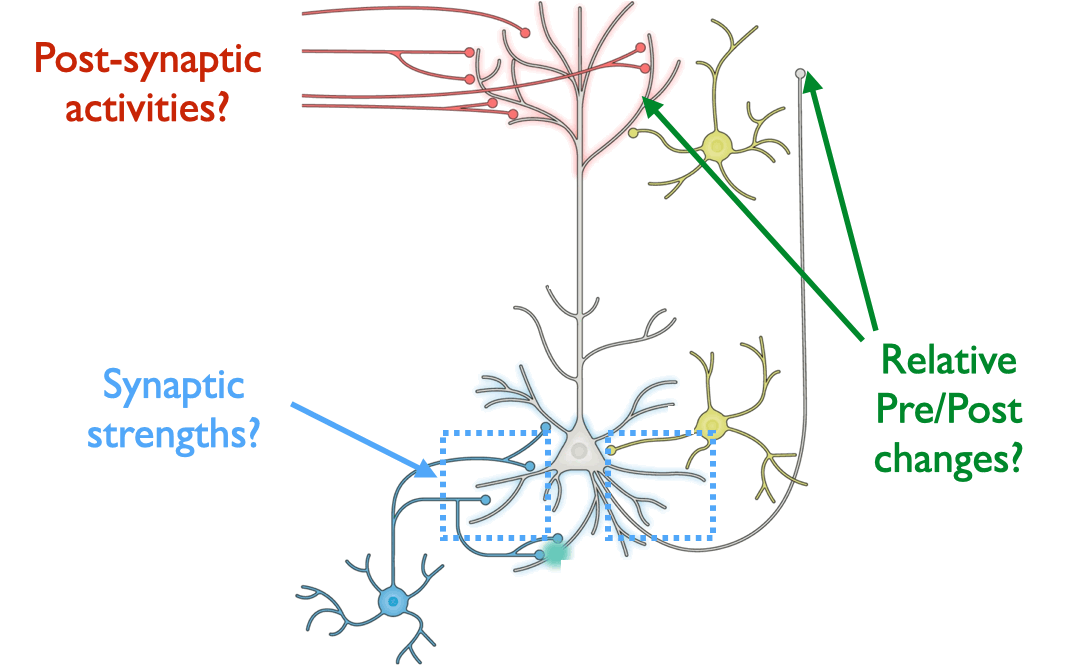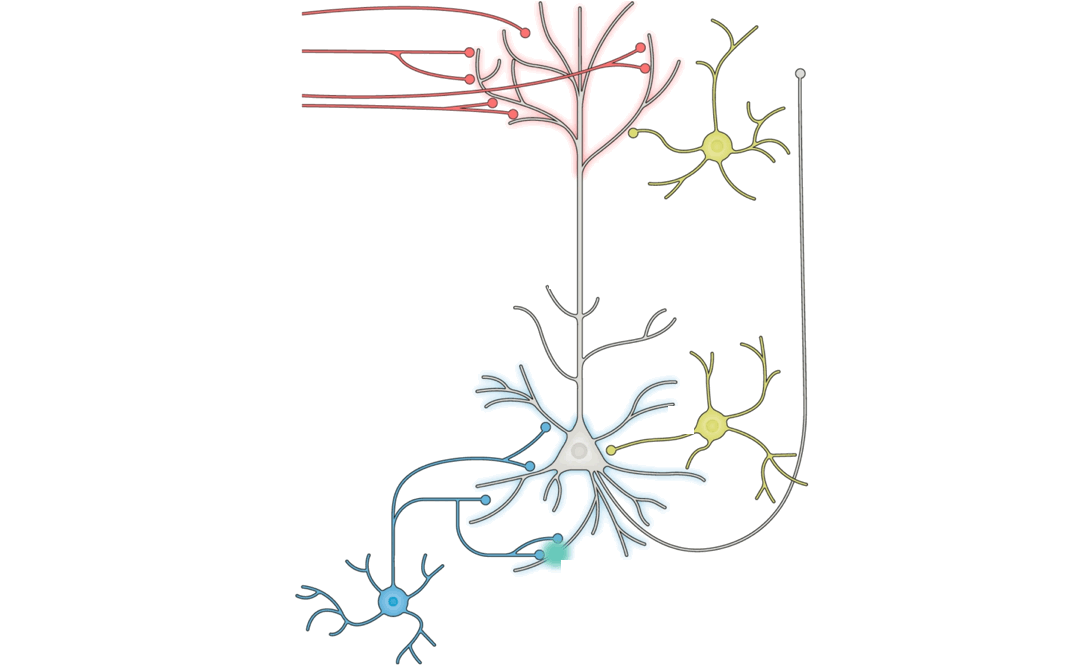
Answering this question turns out to be a substantial challenge, because it is difficult on purely theoretical grounds to identify which patterns of neural changes arise from given learning rules, without also knowing the overall network connectivity and reward target (if any) of the learning system.
But, there may be a silver lining. While ANNs consist of units that are highly simplified with respect to biological neurons, recent progress within the past few years has shown that the internal representations that emerge in trained deep ANNs often overlap strongly with representations in the brain, and are in fact quantifiably similar to many neurophysiological and behavioral observations in animals [7]. For instance, task-optimized, deep convolutional neural networks (CNNs) have emerged as quantitatively accurate models of encoding in primate visual cortex [8, 9, 10]. This is due to (1) their cortically-inspired architecture, a cascade of spatially-tiled linear and nonlinear operations; and (2) their being optimized to perform certain behaviors that animals must perform to survive, such as object recognition [11]. CNNs trained to recognize objects on ImageNet predict neural responses of primate visual cortical neurons better than any other model class. Thus, these models are, at the moment, some of our current best algorithmic “theories” of the brain -- a system that was ultimately not designed by us, but rather the product of millions of years of evolution. On the other hand, ANNs are designed by us -- so the ground truth learning rule is known and every unit (artificial “neuron”) can be measured up to machine precision.
Can we marry what we can measure in neuroscience with what we can conclude from machine learning, in order to identify what experimentally measurable observables may be most useful for inferring the underlying learning rule? If we can’t do this in our models, then it seems very unlikely to be able to do this in the real brain. But if we can do this in principle, then we are in a position to generate predictions as to what data to collect, and whether that is even within reach of current experimental neuroscience tools.
Methods
We adopt a two-stage “virtual experimental” approach. In the first stage, we train ANNs with different learning rules, across a variety of architectures, tasks, and associated hyperparameters. These will serve as our “model organisms” on which we will subsequently perform idealized neuroscience measurements. In the second stage, we calculate aggregated statistics (“measurements”) from each layer of the models as features from which to train simple classifiers that classify the category that a given learning rule belongs to (specified below). These classifiers include the likes of a linear SVM, as well as simple non-linear ones such as a Random Forest and a 1D convolutional two-layer perceptron.
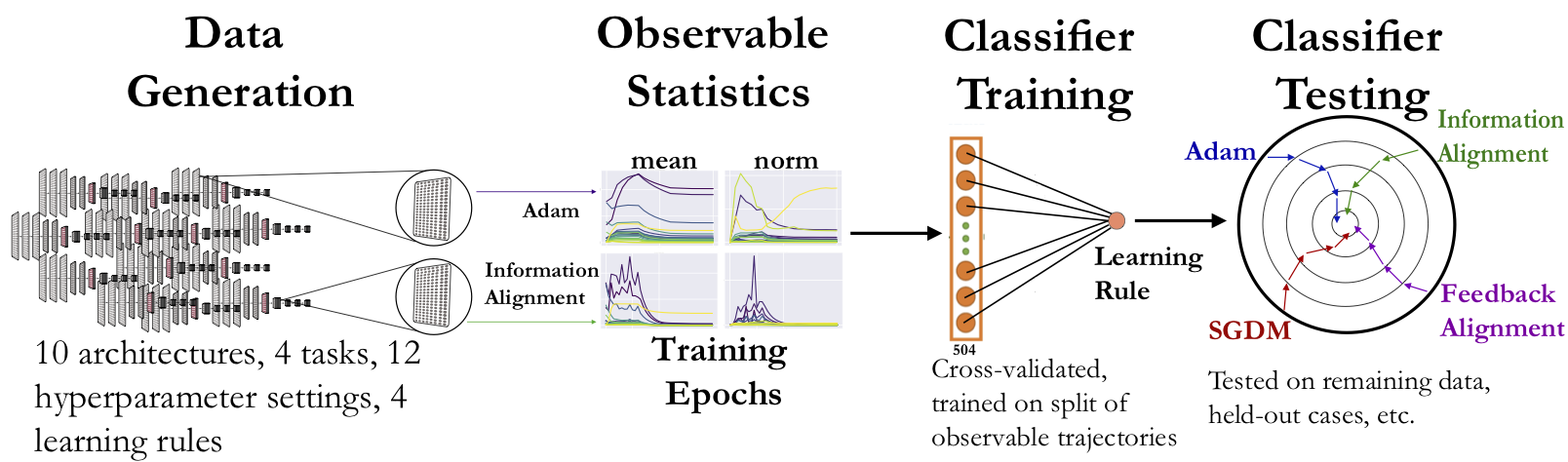
Generating a large-scale dataset is crucial to this endeavor, in order to both emulate a variety of experimental neuroscience scenarios and be able to derive robust conclusions from them. Thus, in the first stage, we train ANNs on tasks and architectures that have been shown to explain variance in neural responses from sensory (visual and auditory) brain areas [8, 12]. These include supervised tasks across vision and audition, as well as self-supervised ones. We consider both shallow and deep feedforward architectures on these tasks, that are of depth comparable to what is considered reasonable from the standpoint of shallower non-primate (e.g. mouse [13]) and deeper primate sensory systems [8, 14, 15].
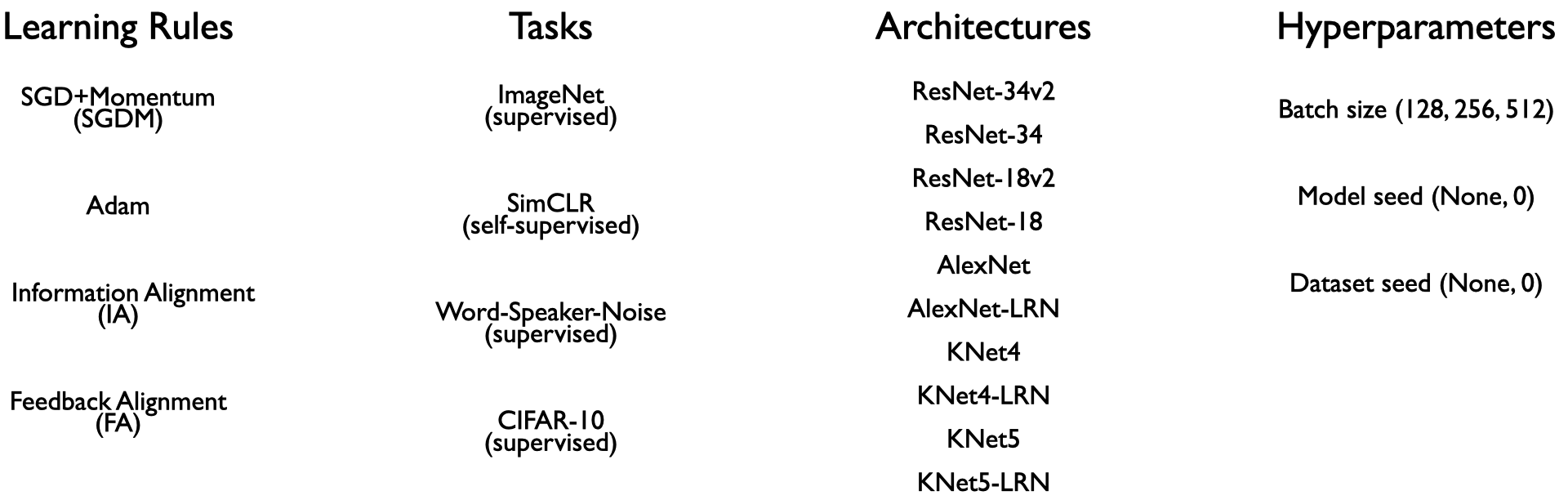
In the second stage, we train classifiers on the observable statistics from these ANNs to predict the learning rules (as specified in the table above) used to train them. The four learning rules were chosen as they span the space of commonly used variants of backpropagation (SGDM and Adam), as well as potentially more biologically-plausible “local” learning rules (Feedback Alignment (FA) and Information Alignment (IA)) that efficiently train networks at scale to varying degrees of performance but avoid exact weight symmetry.
Because the primary aim of this study is to determine the extent that different learning rules led to different encodings within ANNs, we begin by defining representative features that can be drawn from the course of model training. For each layer in a model, we consider three measurements: weights of the layer, activations from the layer, and layer-wise activity change of a given layer’s outputs relative to its inputs. We choose ANN weights to analogize to synaptic strengths in the brain, activations to analogize to post-synaptic firing rates, and layer-wise activity changes to analogize to paired measurements that involve observing the change in post-synaptic activity with respect to changes induced by pre-synaptic input.

For each measure, we consider three functions applied to it: “identity”, “absolute value”, and “square”. Finally, for each function of the weights and activations, we consider seven statistics, and for the layer-wise activity change observable, we only use the mean statistic due to computational restrictions. This results in a total of 45 continuous valued observable statistics for each layer, though 24 observable statistics are ultimately used for training the classifiers, since we remove any statistic that has a divergent value during the course of model training. We also use a ternary indicator of layer position in the model hierarchy: “early”, “middle”, or “deep” (represented as a one-hot categorical variable).
We Can Separate Learning Rules from Aggregate Statistics of the Weights, Activations, or Layer-wise Activity Changes
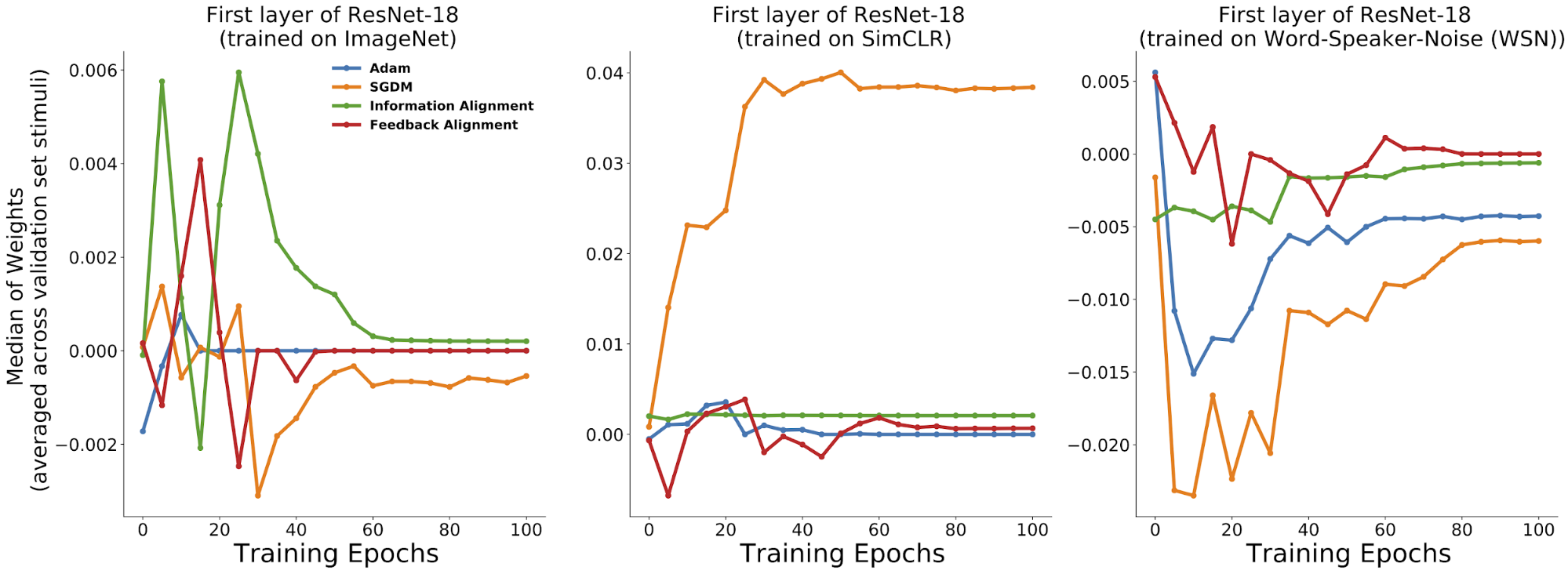
Already by eye, one can pick up distinctive differences across the learning rules for each of the training trajectories of these metrics. Of course, this is not systematic enough to clearly judge one set of observables versus another, but provides some initial assurance that these metrics seem to capture some inherent differences in learning dynamics across rules.
So these initial observations seem promising, but we want to make this approach more quantitative. Suppose for each layer we concatenate the trajectories of each observable and the position in the model hierarchy that this observable came from. Can we generalize well across held-out examples?
It turns out that the answer is in fact, yes. Across all classes of observables, the Random Forest attains the highest test accuracy, and all observable measures perform similarly under this classifier.
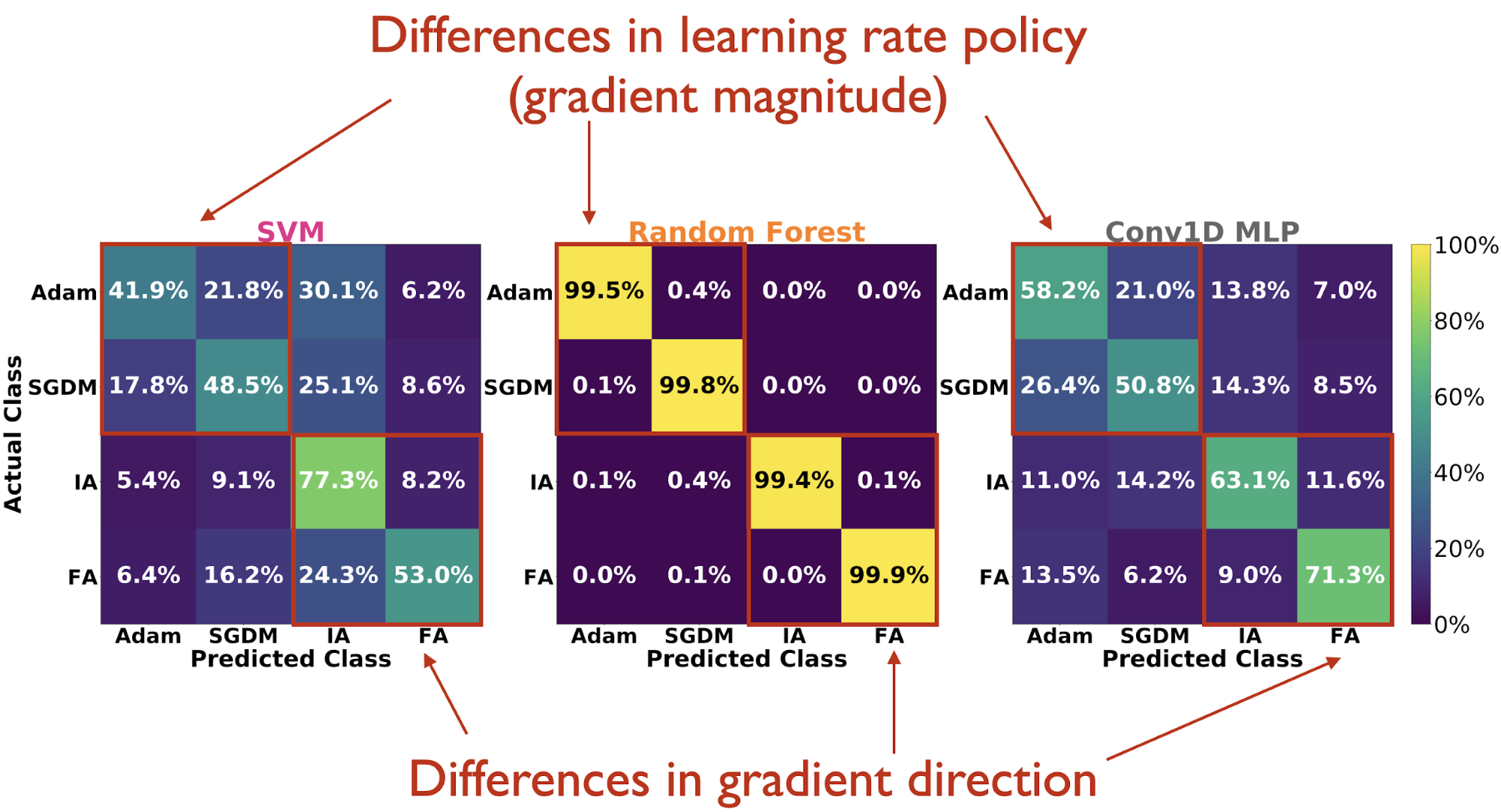
Looking at confusion matrices on the test set, we see that the Random Forest hardly mistakes one learning rule from any of the others. And when the classifiers do make mistakes, they generally tend to confuse Adam vs. SGDM more so than IA vs. FA, suggesting that they are able to pick up more on differences (reflected in the observable statistics) due to high-dimensional direction of the gradient tensor than the magnitude of the gradient tensor (the latter being directly tied to learning rate policy).
Adding Back Some Experimental Neuroscience Realism
Up until this point, we have had access to all input types, the full learning trajectory, and noiseless access to all units when making our virtual measurements of ANN observable statistics. But in a real experiment where someone were to collect such data from a neural circuit, the situation would be far from this ideal scenario. We therefore explore experimental realism in several ways, in order to identify which observable measures are robust across these scenarios.
Access to only portions of the learning trajectory: subsampling observable trajectories
The results presented thus far were obtained with access to the entire learning trajectory of each model. Often however, an experimentalist collects data throughout learning at regularly spaced intervals. We capture this variability by randomly sampling a fixed number of points at a fixed temporal spacing for each trajectory, which we refer to as a “subsample period”.
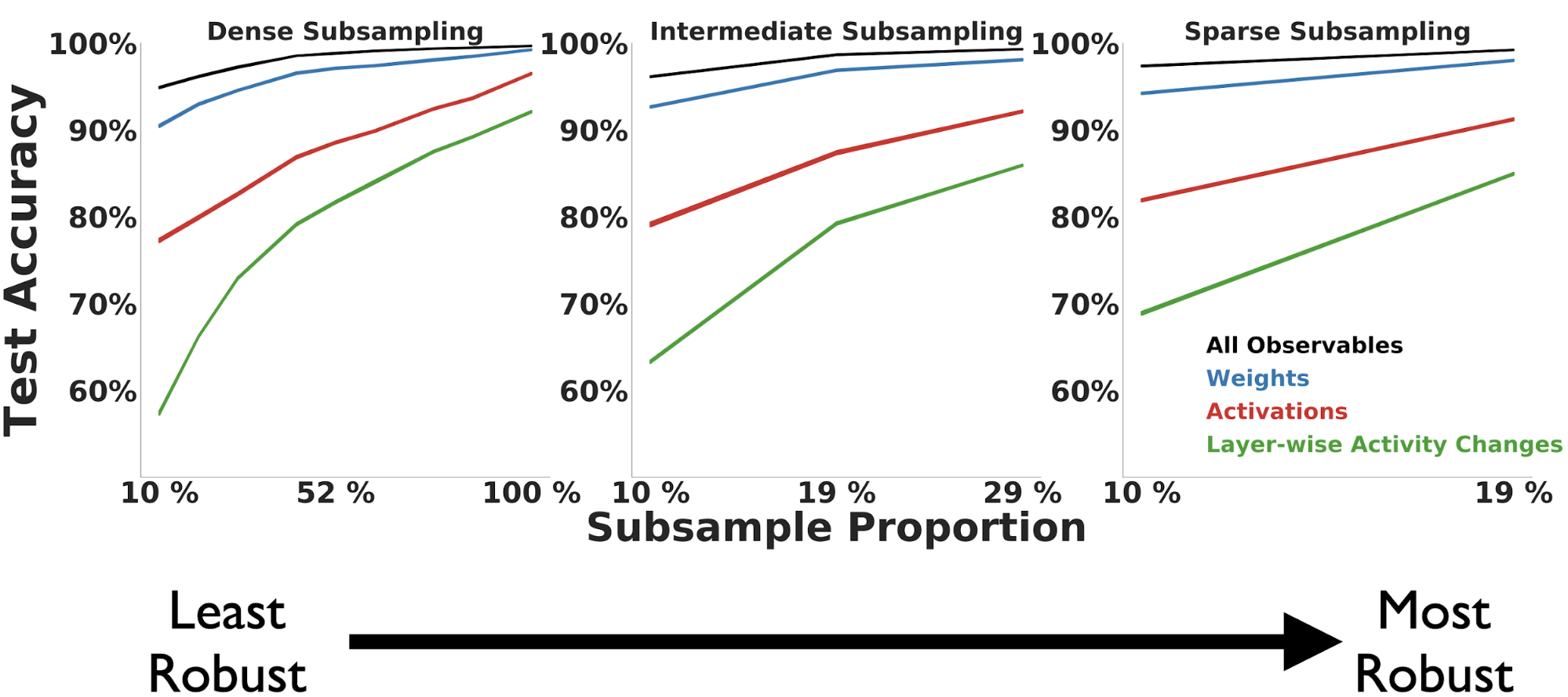
We find across observable measures that robustness to undersampling of the trajectory is largely dependent on the subsample period length. As the subsample period length increases (in the middle and right-most columns), the Random Forest classification performance increases compared to the same number of sampled points for a smaller period (depicted in the left-most column).
Taken together, these results suggest that data consisting of measurements collected temporally further apart across the learning trajectory is more robust to undersampling than data collected closer together in training time. Furthermore, across individual observable measures, the weights are overall the most robust to undersampling of the trajectory, but with enough frequency of samples we can achieve comparable performance with the activations.
Incomplete and noisy measurements: subsampling units and Gaussian noise before collecting observables
The aggregate statistics computed from the observable measures thus far have operated under the idealistic assumption of noiseless access to every unit in the model. However, in most datasets, there is a significant amount of unit undersampling as well as non-zero measurement noise. How do these two factors affect learning rule identification, and in particular, how noise and subsample-robust are particular observable measures?
Addressing this question would provide insight into the types of experimental neuroscience paradigms that may be most useful for identifying learning rules, and predict how certain experimental tools may fall short for given observables. For instance, optical imaging techniques can use fluorescent indicators of electrical activities of neurons to give us simultaneous access to thousands of neurons. But these techniques can have lower temporal resolution and signal-to-noise than electrophysiological recordings that more directly measure the electrical activities of neurons, which in turn may lack the same coverage.
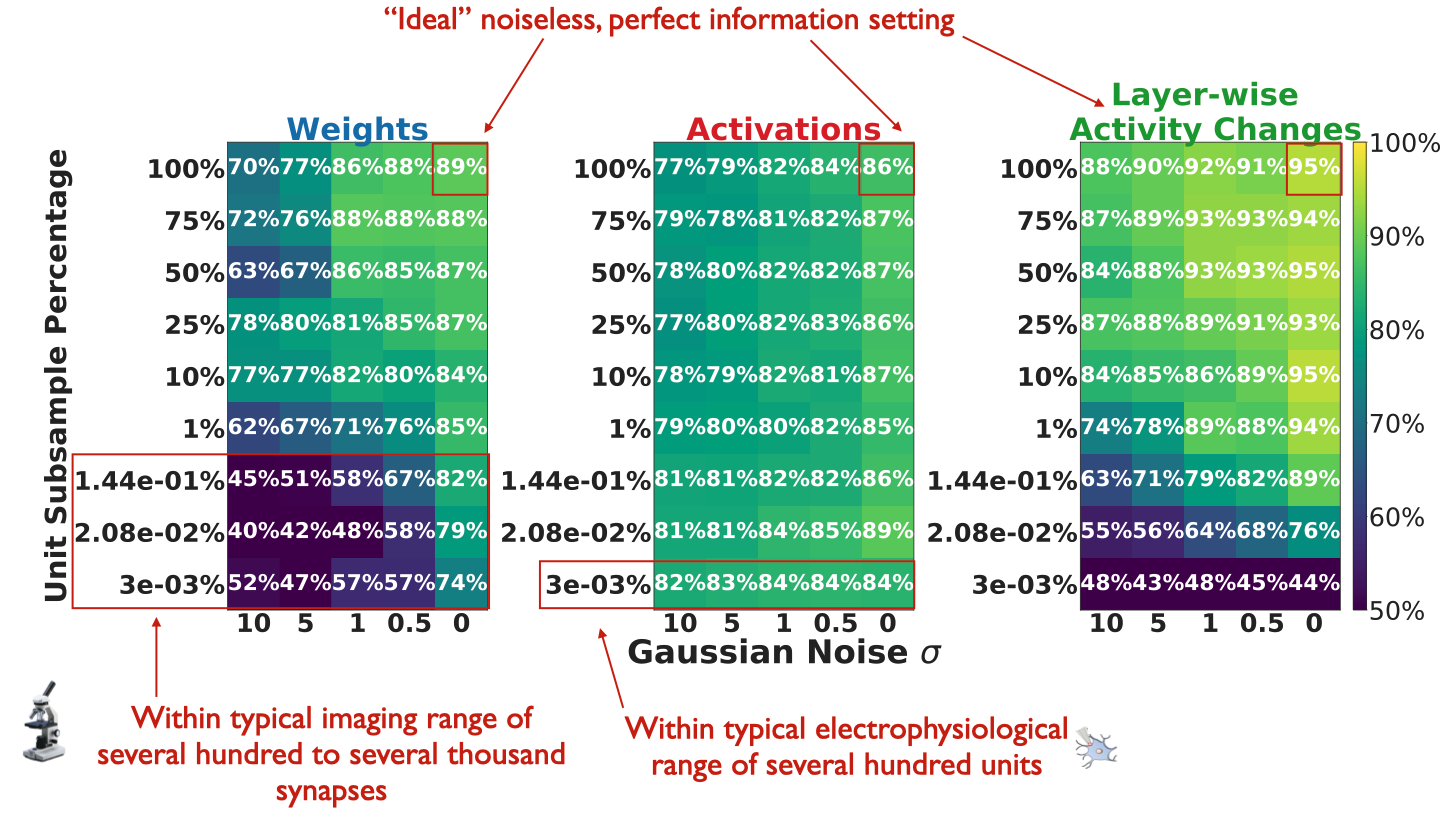
To account for these tradeoffs, we model measurement noise as an additive white Gaussian noise process added to units of ResNet-18 trained on the ImageNet and self-supervised SimCLR tasks. We choose IA vs. FA since the differences between them are conceptually stark: IA imposes dynamics on the feedback error weights during learning, whereas FA keeps them fixed. If there are scenarios of measurement noise and unit subsampling where we are at chance accuracy for this problem (50%), then it may establish a strong constraint on learning rule separability more generally.
Our results suggest that if one makes experimental measurements by imaging synaptic strengths, it is still crucial that the optical imaging readout not be very noisy, since even with the amount of units typically recorded currently (on the order of several hundred to several thousand synapses), a noisy imaging strategy of synaptic strengths may be rendered ineffective.
Instead, current electrophysiological techniques that measure the activities from hundreds of units could form a good set of neural data to separate learning rules. Recording more units with these techniques can improve learning rule separability from the activities, but it does not seem necessary, at least in this setting, to record a majority of units to perform this separation effectively.
Conclusions
As experimental techniques in neuroscience continue to advance, we will be able to record data from more neurons with higher temporal resolution. But even if we had the perfect measurement tools, it is not clear ahead of time what should be measured in order to identify the learning rule(s) operative within a given neural circuit, or whether this is even possible in principle. Our model-based approach demonstrates that we can identify learning rules solely on the basis of standard types of experimental neuroscience measurements from the weights, activations, or layer-wise activity changes, without knowledge of the architecture or loss target of the learning system.
Additionally, our results suggest the following prescription for the type of experimental neuroscience data to be collected towards this goal:
Electrophysiological recordings of post-synaptic activities from a neural circuit on the order of several hundred units, frequently measured at wider intervals during the course of learning, may provide a good basis on which to identify learning rules.
We have made our dataset, code, and interactive tutorial publicly available so that others can analyze these properties without needing to train neural networks themselves. Our dataset may also be of interest to researchers theoretically or empirically investigating learning in deep neural networks. For further details, check out our NeurIPS 2020 paper.
Acknowledgements
I would like to thank my collaborator Sanjana Srivastava and advisors Surya Ganguli and Daniel Yamins. I would also like to thank Jacob Schreiber, Sidd Karamcheti, and Andrey Kurenkov for their editorial suggestions on this post.